ABSTRACT
Evidence is presented for existing and hypothetical structures and functions which 'bridge the explanatory gap' to mechanistically explain how individual consciousnesses could be derived from a higher (God) consciousness through an interface created in the brain by endogenous (emitted from within) light. It is hypothesized that photons emitted from cells in the brain are guided to the surfaces of the brain's fluid-filled ventricular spaces, where the photons interact with beating cilia lining those ventricles and are guided by the beat timing to form interference patterns in the ventricular spaces, creating an interface (a nexus) through which a tiny portion of the "light of God" is able to animate the corpus. Some of the necessary mechanisms such as light emissions from cells are known; others are hypothetical. Fortunately some of the hypothesized mechanisms of this model of consciousness are readily testable.
INTRODUCTION
Endogenous light refers to cell photon emissions that in this model are physiologically functional. Not to be confused with macroscopic bioluminescence, cell photon emissions occur at low intensities ('weak' or 'ultraweak' they may be called) not detectable by the eye. They span the visible spectrum and may extend into the near ultraviolet and infrared; although emissions outside the visible spectrum may only loosely be termed 'light', they are included in that term in this context. For a system of physiologically functional endogenous light to exist, at least four capabilities are required (though others beyond these minimal four may also exist):
- Light signals must be generated and emitted from cells.
- Emitted light signals must be transmitted to adjacent and/or distant cells.
- Cells must be capable of receiving light signals.
- Cells must be capable of transducing received light signals into information they can further process.
The first criterion, cell photon emission, is well documented but has not yet been shown to carry information except for some examples of macroscopic bioluminescence, fireflies being perhaps the best known. The second criterion may be met by collagen fibers functioning as fiber optics, since almost all nonmotile cells are interconnected by a network of collagen fibers, a network which makes up approximately half of the total protein content of a human body. Other fiber types such as elastin and intracellular matrixes of microtubules (especially those running the length of neural axons) are also possible candidates for functional roles in a biophotic communication network. Collagen fibers have been shown to exhibit fiber optic properties in preferentially conducting light along their fiber axes, and collagen fibers will even modulate a light signal passing through them when stretched at physiologic force levels. It is well known that retinal photoreceptors are capable of meeting criterion 3 (receiving light) and 4 (processing light into information); it is proposed that retinal photoreception represents a functional specialization for reception of light in an evolutionary sense in the same way that macroscopic bioluminescence is a functional specialization for light emission: both specializations must be built upon more rudimentary capabilities of relatively primitive cells. In that context two facts are significant: the first is that the complex lamellar structures of retinal photoreceptors are highly specialized cilia, and the second is that ciliated cells are abundant in the brain and configured in such a way that they could dynamically exchange endogenous photons across the ventricular spaces. Do cilia possess rudimentary photoreceptive (and possibly waveguide) capabilities, which evolution has built upon to create photoreceptors? This is a key question because much of the innermost ependymal lining of the cerebral ventricles is ciliated (with the cilia having no known function). It is proposed that ventricular ciliary beating is influenced by and becomes coordinated with the timing of neural activity so that endogenous light emissions are stimulated and guided out of ventricular cilia into dynamically resonant patterns in the ventricles and possibly some surrounding tissues in a process termed neurophotic resonance. Reception of endogenous photons by cilia and other structures such as the pineal organ would in turn influence neural events as part of the feedback process needed to maintain neurophotic resonance. These dynamically resonant patterns of interacting light would involve energy loci where photon fluxes could influence each other and thereby gain feedback capability on neural events. The dynamic photonic structure so formed in the brain's spaces therefore becomes a nexus able to connect consciousness to the body. That nexus of endogenous light occupies a certain volume, up to about 150 ml ventricular space in a human brain although it may not all be used. It is generated by the mechanisms for neurophotic resonance but is not itself conscious, it is a receiver for consciousness. That's because the volume of the nexus is permeated by another source of light which is able to interact with the endogenous light of the nexus and influence it, thus affecting the entire neurophotic resonance. The other source of light available to interact with endogenous photons is the larger population of virtual photons filling space outside our normal frame of reference. Conventional wisdom has virtual photons popping in and out of existence even within our own frame of reference, but has little to say about the vast network of interconnected light that virtual photons must inhabit outside of our normal frame of reference because light experiences no time. The endogenous light nexus thus brings a tiny volume of that virtual photon network into contact with and control of a brain and body, and it is in that virtual network that the self-awareness of consciousness actually resides. Only a small portion of it, a 'bleb' is able to be expressed within the relatively tiny volume of a nexus though, in the same way that a very large hologram when cut into small portions retains a rough image of the original on each portion but lacks a lot of detail. The whole is out there as a vast network of interconnected light simultaneously traversing all of space and time, where resides the unity of intelligent consciousness underlying the universe which in simplistic historical terminology many would term 'God'. It is not an external God though, if God is even the right term, because we are ourselves occupied by tiny portions at a time which we experience as personal consciousness, that elusive self-awareness. We can think of our individual conscsciousnesses as blebs of God in the plainest sense, like drops from an ocean of consciousness much greater than we can ever hope to fathom. Those blebs are supported in our brains by the endogenous light nexus creating a pathway for a small part of the intelligent consciousness underlying the universe to interact with and influence the dynamic patterning of the nexus, which in turn influences neural events, which in turn influence perception, thought, and behavior in resonant feedback loops.
ENDOGENOUS LIGHT SOLVES THE HARD PROBLEM
Light is an ancient and pervasive spiritual metaphor
|
|
Some people who have near-death experiences (NDEs) report encounters with a Clear Light or a White Light they identify as God. On the basis of spiritual tradition and NDE anecdotal evidence it is hypothesized (1) that God exists, is intelligently self-aware, and is somehow formed of some kind of light.
Relativistic Frames of Reference
Light is curious stuff, baffling to us in its wave-particle duality and relativistic properties. Time does not exist for light. It may take centuries to travel from a distant star to fall on the retina of an observer's eye, but as far as the light is concerned it exists continuously along its path, in its frame of reference it is not at one place at one time and farther along at another time. Only for matter and to our matter-bound consciousness is there a time-bounded frame of reference. All of spacetime is therefore crisscrossed by light from various sources which, in light's frame of reference, is all present and crisscrossing at once. A holographic model is applicable to this view (Talbot, 1992), with constructive and destructive interference at the crisscrossings creating a complex design throughout the universe. How time enters the picture is a great mystery, but physicists speak of virtual photons (and virtual particles) pervading spacetime such that even a tiny point in a hard vacuum contains an enormous amount of energy. It appears that our time-bounded frame of reference allows us to perceive only one very thin 'slice at a time' of light's frame of reference. Hypothesis 2: God-intelligence is a property of all the light of the universe existing simultaneously in its own frame of reference and forming a vastly complex holographic interference pattern throughout spacetime. The 'all at once' quality of its simultaneity is sometimes experienced (when we notice it) as a characteristic feature in each time-slice of our individual consciousness.
Blebs of God
Divide a hologram in half, and each piece still contains the entire holographic image. Repeat the process many times and each piece still contains the whole image, but gradually the resolution of the image degrades and fine detail disappears. Eventually only a very poor rendition of the original remains.
Now imagine isolating a tiny bleb of light's holographic frame of reference so that it is distinct from the rest of the vast holographic web of God-intelligence. The original is not noticeably changed, but a very poor version of the original is on the bleb. We'll call it a 'Bleb of God', having these properties:
- It's made of the same stuff as the original.
- It's got the whole image on it, although dimly and missing details.
Hypothesis 3: we are conscious beings because we are Blebs of God. We have self-aware experiences of sensation, thought, and feeling because these are properties of the God-intelligence underlying the universe, and tiny blebs of that intelligence are able to inhabit physical matter of the proper construction (brains). Blebs of God are formed from the same light as God but occupy only a tiny spacetime volume. Therefore they hold a very poor rendition of God-intelligence, and although they may have a complete representation of the overall holographic pattern, they lack most of the original's detail.
Hypothesis 4: Blebs of God Interfere With Endogenous Light in the Brain
In light's timeless frame of reference, a volume of our time-bounded spacetime containing dynamic holographic patterns of endogenous light within a brain overlaps all the other light that ever did or ever will cross that volume. That volume of spacetime is the volume from which a Bleb of God is formed; because it overlaps the holographic optical element formed from endogenous light, the two holographic patterns may interfere and thereby dynamically modify the interaction of endogenous light with the neural circuits of the brain. This has several effects:
- The Bleb of God interfaces with and gains control of resonant neural circuits.
- The Bleb of God receives and interprets information returned from resonant neural circuits.
- The Bleb of God animates the corpus (otherwise a zombie) with its dim God-intelligence, which we know as consciousness.
- The Bleb of God's intelligent perception is dominated by sequential slices of time-bounded endogenous light inputs because they have higher resolution and contain more detail.
- Only those aspects of the Bleb of God's potential for which there are corresponding neural resonant circuits in the brain are expressed.
ENDOGENOUS LIGHT IN THE BRAIN
It is proposed that ventricular ependymal cilia function as optical waveguides to trap and guide endogenous photons into and out of ependymal cells across ventricular spaces. A photon captured by a cilium is guided by the ciliary microtubules into the ependymal cell and then reflected by the centrioles at the base of the cilium into any one of many possible cytoskeletal pathways within the cell. Photons may possibly be conducted through the cytoskeletal microtubules (Jibu et al., 1994) or absorbed by them so that the information they carry subsequently propagates as a series of quantum events moving along microtubular subunits in the manner described by the Penrose-Hameroff 'Orch OR' model (Hameroff, 1998).
In this model, photon energy absorbed by ependymal cells can have many possible destinations within those cells and thereby multiple possible effects. Absorption of a photon along the cilium or very near its base may affect the dynamics of ciliary beating (i.e., beat frequency and the swept field). Absorption of a photon by a centriole may ratchet the centriole's orientation so that subsequent photons get aimed at different cytoskeletal pathways. If photons are not absorbed until they reach mitochondria, they may cause the stimulated emission of more endogenous photons. The transmission of a photon's energy to the cell nucleus may influence gene expression. The destination of any given photon may be governed by properties of the photon itself, such as frequency, phase, and polarization (plane and circular). However, in the present context the issue is how endogenous photons may control the resonance of neural circuits:
Control of Neural Resonance
Microtubular quantum events (Hameroff, 1998) or possibly microtubular electron currents (Insinna, in press) may be transmitted to neurons through desmosomal connections with ependymal cells or possibly through very localized changes in ependymal cell resting membrane potentials which then affect the resting potentials of adjacent neuronal dendrites. The end result in either case is that absorbed photons influence the membrane thresholds of 'critical neurons'. This affects the generation of action potentials in specific resonant circuits that may be increased in firing probability through constructive interference or decreased in firing probability through destructive interference that is largely dependent upon the timing of photon absorption at the ependymal cell and the moment-to-moment distribution of membrane potentials at neurons influenced by ependymal cell photon absorption. The resonance of any neural circuit is therefore dependent upon the synchrony of all electrical inputs arriving at specific loci on those critical neurons and is ultimately governed by the timing of photic signals arriving at their associated ependymal cells. Like many naturally resonant systems, only a relatively small driving input is required to conrol the amplitude of the resonance and to shift its phase.
Photon Emission
Endogenous photons measured in the laboratory appear to originate randomly from mitochondria, but their emissions are known to be stimulated at discrete intervals or in response to external stimuli by some bioluminescent organisms. It is likely that both spontaneous and stimulated emissions contribute to the ventricular light environment, with stimulated emissions dominating the induction of neural resonance (the timing of stimulated emissions would of course be crucial to the timing of received photons). As described above, endogenous photons conducted through the cell cytoskeleton to mitochondria may themselves stimulate the emission of more endogenous photons from mitochondria and in this manner maintain a sufficiently high population of properly timed intraventricular photons to drive neural resonance.
Figure 1: Lateral View of the Ventricles (fluid-filled spaces)
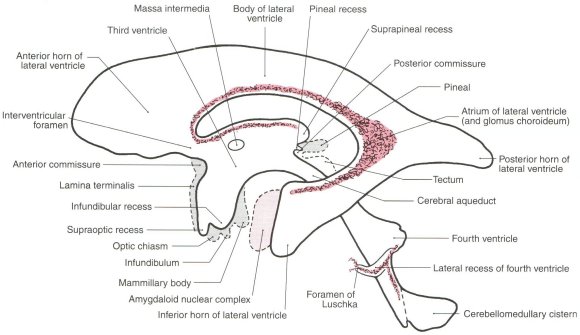
Figure 2: Dorsal View of the Ventricles
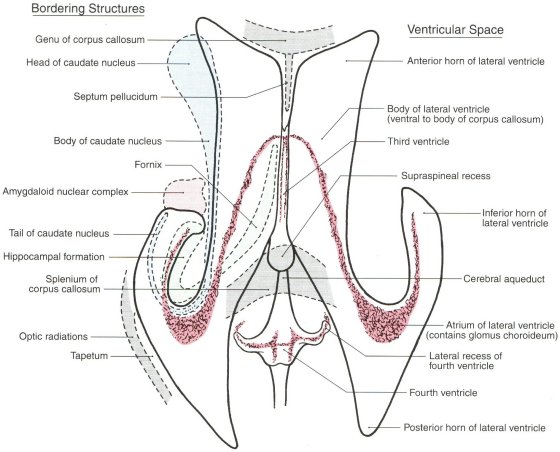
Adapted from Haines, D.E. Neuroanatomy: An atlas of structures, sections, and systems, 5th ed. Lippincott Williams & Wilkins, Baltimore, 2000
Cilia occupy much of the ependymal surfaces lining the brain's ventricles; their function there is unknown. It is proposed that they function to capture and guide endogenous photons that are transmitted across the ventricular spaces from opposing ependymal surfaces, and that they guide the direction in which photons are emitted from ependymal cells into the ventricular spaces.
Figure 3: Ventricular Cilia as Waveguides

Ventricle surface of rabbit, largely lined by ciliated ependymal cells. Ci = cilia; SC = supraependymal cells; Ta = tanycytes.
DEVELOPMENT
The cavity of the embryonic neural tube develops into the brain's ventricles. As the brain develops, a layer of cells on the inner walls of the neural tube gives rise to all (or almost all) of the brain's neurons, which each migrate outward along glial cell processes to find their correct position and functional relationships with multitudes of other neurons. The innermost layer of cells is primarily ciliated ependymal cells. Endogenous light supporting consciousness in the ventricles is consistent with this original relationship (ventricular ependymal layer --» cortex) being retained in the adult brain.
Figure 4: Ventricular-centric development
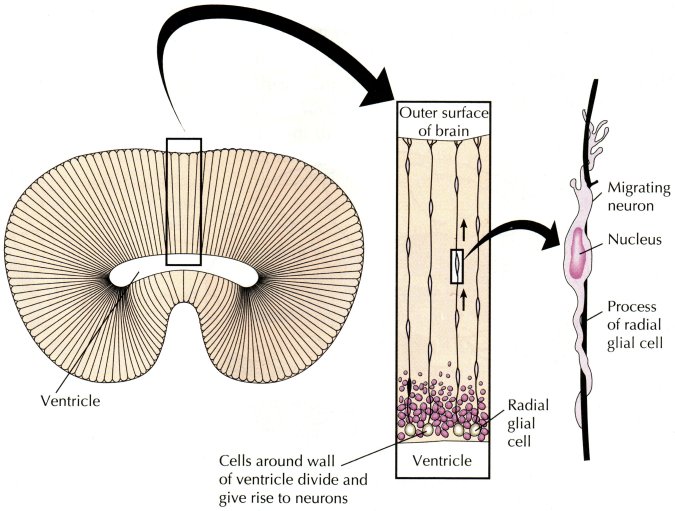
Adapted from Carlson, N.R. Physiology of Behavior, 5th ed. Allyn and Bacon, Boston, 1994 p. 84, where it was first adapted from Bloom, F.E., and Lazerson, A. Brain, Mind, and Behavior, 2nd ed. W.H. Freeman, New York, 1988.
The Central Role of the Thalamus
More Than a Relay Station
All sensory inputs except olfaction first project to thalamic nuclei before they are relayed to the cortex. Yet the number of these projections is minor compared to the many more corticothalamic projections from cortical association areas (Newman, 1995), and the number of reciprocal corticothalamic projections is an order of magnitude greater than the number of thalamocortical projections (Llinas and Pare, 1991). The nucleus reticularis, an outer 'shell' of the thalamus, appears to control the gating of neural resonant traffic between thalamus and cortex, being "the primary generator of widespread synchrony in the EEG" (Newman, 1995).
Thalamic-Ventricular Relationships and Neural Resonance
The thalamus is a double organ divided by the third ventricle and bounded along much of its dorsal surface by the lateral ventricles. It is proposed that endogenous light within the third and lateral ventricles controls the neural activity of thalamic nuclei having projections to the nucleus reticularis gating circuits, to regulate the flow of information returned from the cortex. Newman's thalamocortical map below illustrates the precise mapping between thalamus and cortex that is largely reciprocated in corticothalamic connections as well. Note the convergence of thalamic areas mapping to prefrontal cortex (where higher-level decision-making is believed to be processed) at the medialdorsal nuclei of the two thalami (MD, PfC) which lie opposite one another across the space of the third ventricle. Neural resonance between prefrontal cortex and the medialdorsal nuclei would in this model be regulated by endogenous light in that space, controlling relevant nucleus reticularis gates between cortex and thalamus. In this model the raw sensory information which first converges at the thalamus and endogenous light nexus in the ventricles of the brain is also passed through to cortex which then returns interpreted information about the sensory inputs back to consciousness in the ventricles in a constant process of input analysis.
Figure 5: Newman's Thalamo-Cortical Map
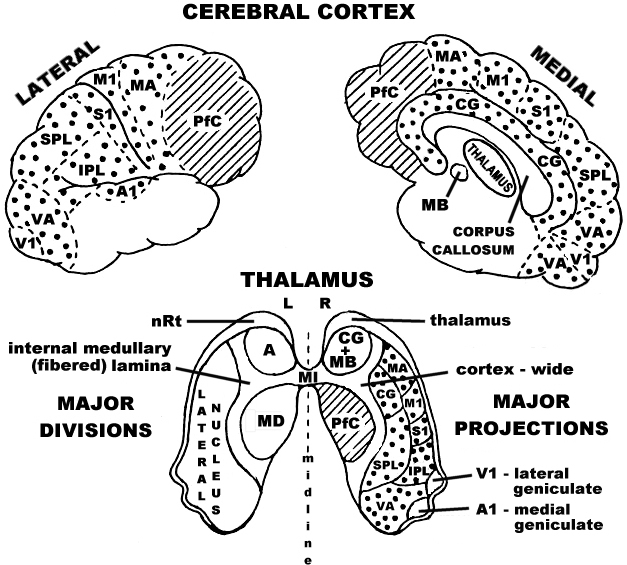
The major divisions of the thalamus and the major projections it shares with the cerebral cortex. (Adapted from Newman, 1995).
|
|
ANALOGOUS OPTICAL COMPUTING MODEL
In this design for an optical supercomputer, numerous laser diode signal sources send photons through space to pass information to a holographic optical element which directs the light signals to desired memory-cell locations. Gate circuits called optical input gates (consisting of an optical detector and an inverter) transform the light signals to electrical signals usable by the memory cells. A similar design and prototype has been developed for an optical central processing unit. The major difference between this design and a ventricular optical processing system is that the holographic optical element depicted below is a reflective hologram that directs light back to receiving elements on the same plane as the emitting laser diodes; in a ventricular optical system the holographic element formed by endogenous light would be a transmission hologram that would modulate the light passing from one ventricular surface to another.
Figure 7: Holographic Computing
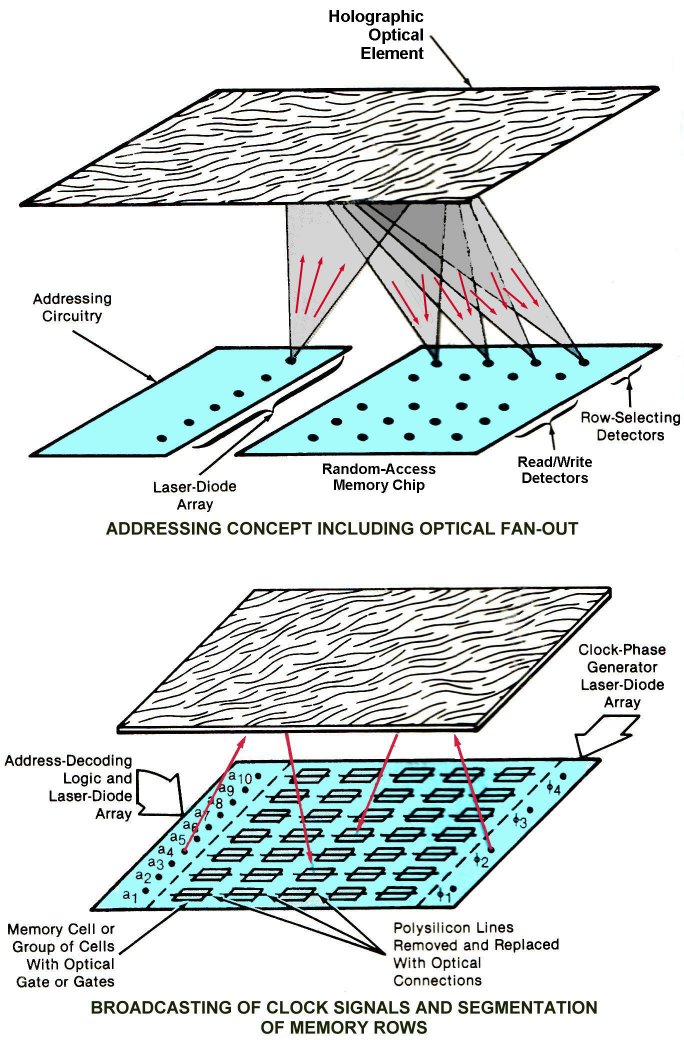
Adapted from Johnson AR, Nixon RH, and Bergman LA. Optical Addressing and Clocking of RAM's. NASA Tech Briefs 13:32-34, 1989. See also Wu et al., 1987.
HOLOGRAPHIC OPTICAL ELEMENT
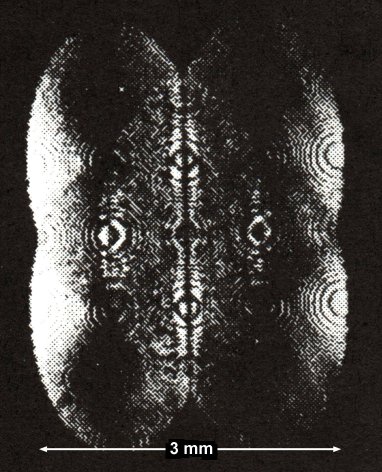
Figure 8: Actual Holographic Optical Element
|
An actual holographic optical element used in an optical computer prototype. Computer-generated holographic optical element printed on silicon using methods compatible with existing integrated circuit manufacturing. Reflectivity was increased by evaporating aluminum onto the surface. The holographic optical element defines and implements the interconnection patterns between sources and detectors. Using a holographic optical element rather than electrical connections or optical fibers allows greater freedom in designing interconnections and in reprogramming if necessary. A ventricular holographic element might somewhat resemble this interference pattern but would be dynamic instead of static and probably multicolored. Adapted from Wu et al., 1987. |
CELL PHOTON EMISSION
Evolutionary Perspective
Bioluminescence and ultraweak cell photon emissions are well documented phenomena, with origins in the most primitive organisms. Bacterial bioluminescence occurs in some species with such intensity that their light emissions are visible to the naked eye; bacteria may commit nearly five percent of their soluble protein to luciferase, an enzyme catalyzing light emission. Thus there is a considerable metabolic energy expenditure in the synthesis of luciferase, then further energy is expended as photons, but the function of bacterial bioluminescence remains unknown. In evolutionary terms such energy expenditures are generally associated with some selective advantage; whatever those advantages might be, it is reasonable to expect bacterial capabilities to be retained and perhaps elaborated upon by higher organisms. In fact very diverse organisms such as dinoflagellates, coelenterates, squid, shrimp, jellyfish, fireflies, and some fishes emit light using luciferase or similar mechanisms. Their light emissions occur on a macroscopic scale detectable by the human eye, and originate from protein-enzyme systems like the luciferin and luciferase of bacteria. In the case of fireflies and fishes, their light emissions are known to serve communicative purposes, though their photon exchanges represent inter-organism communication and not inter-cellular communication. It is likely that such highly specialized mechanisms for information transfer evolved from more primitive forms of light emission.
Ubiquity
With sensitive equipment, photon emissions can be measured from nearly all cells. Colli et al. (1955) reported weak visible-range bioluminescence from seedlings of wheat, beans, lentils, and corn. Veselovskii et al. (1963) observed photon emissions from seven to ten day old barley seedlings (Fig. 1). Ruth and Popp (1976) measured photon emissions from yeasts as well as potato and cucumber seedlings. Tarusov et al. (1961) reported photon emissions from liver and muscle tissues of mice. Vladimirov and L'vova (1964) detected emissions from isolated mitochondria of rat liver that occurred most intensely when conditions were optimum for oxidative phosphorylation. Zhuravlev et al.(1973) worked with isolated rat liver mitochondria, finding that mitochondrial luminescence requires ADP and oxygen, and that uncoupled electron transport (non-phosphorylating) can contribute to the observed luminescence. Since photons are emitted when electrons jump from molecular excited states to lower energy levels, it should not be too surprising to find that photons are emitted from mitochondria, where oxidative phosphorylation and electron transport can provide the energetic pathways to boost electrons into excited states. Unlike bacteria and other organisms displaying macroscopic bioluminescence however, precise mechanisms for weak eukaryotic photon emissions remain to be discovered. It has recently been reported that plant cells may emit laser-stimulated emissions (lasing) under certain experimental conditions (Drollett, 2000), for example, but it is unknown if that occurs in vivo. However, it is assumed in the present context that weak photon emissions probably occur from most cell types, including those in the brain.
PHOTON TRANSMISSION
Fiber Optics (intercellular and intracellular)
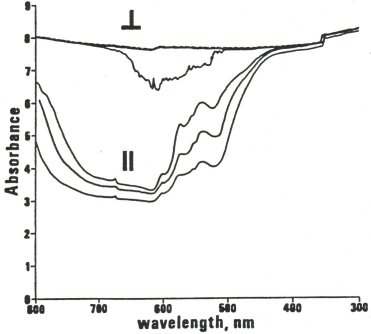
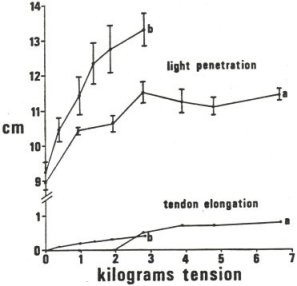
About half of the protein composing a human body is collagen, which forms orderly arrays of fibrillar structures that interconnect and anchor almost all nonmotile cells of the body. However, collagen is more than just scaffolding. For most cells, intact collagen connections seem to be required for them to recognize and express a proper morphological and functional relationship with their neighbors; if cell connections to collagen are disrupted in tissue culture (such as by adding antibodies to the cell adhesion protein fibronectin, breaking those connections), normal cells will rapidly take on the appearance of tumor cells (Yamada et al., 1980). It therefore appears that cell collagen interconnections serve a communicative function too. If the low-level photons that cells emit from their mitochondria do convey information, it seems likely that collagenic fiber optic bundles are part of the medium by which that information is conveyed, because collagen exhibits some fiber optic properties (Simanonok, 1983). A far higher axial conduction of light (more light transmitted parallel to the fibers than perpendicular to them, see figure) is characteristic of fiber optic bundles. However, other fibrous proteins such as elastin (and structures such as axons) may also have roles in intercellular photon transmission.
Figures 9 and 10: Optical properties of collagen bundles
|
|
|
Tendon Fiber Light Absorbance
Spectrophotometry of three 1 cm tendon samples with fibers perpendicular (top; one is dashed and overlays another) and parallel (lower) to the light path. Adapted from Simanonok, 1983. |
Elastooptic light penetration into tendon
Change in light-tendon interaction with applied tension shown in two tendons, a and b. The increase in light penetration with tension is much greater than the tendon elongation, shown at bottom. He-Ne laser at 632.8 nm; error bars represent ďż˝ SD. Adapted from Simanonok, 1983. |
Tracing the extracellular paths of collagen fibers, their connections to cells are through membrane structures called hemidesmosomes, which interface with the cell cytoskeleton. Intracellularly, a finer network of fibers (a complex array of microtubules, microfilaments, and various associated molecules) then extends throughout the cell from those hemidesmosomal connectors. Where and how a photonic signal might be transduced into information is unclear, but one likely destination for intercellular information is DNA, both nuclear and extranuclear. Cell cytoskeletal fibers have been observed extending to mitochondria and cell nuclei, to desmosomes joining adjacent cells, to other hemidesmosomes where cells attach to collagen fibers in the basement membrane, and to ciliary "rootlets" which extend inward from the centrioles at ciliary bases. The cell cytoskeleton therefore may form an intracellular fiber optic system (very small diameter fibers are termed optical waveguides). Selective photon conduction and absorption may occur at various junctures in the cytoskeleton.
Free-Space Photon Transmission
Another potential mechanism for transmitting biophotic signals between cells is across free spaces. An important advantage of this method could be significantly greater system flexibility because the exchange of information in the form of photons would not be hard-wired. A similar method of free-space optical interconnection has been developed for use in optical supercomputers (Wu et al., 1987). As this is an important part of the proposed mechanism for the support of consciousness, it is covered in its own section, Control of Neural Resonance, above.
PHOTON RECEPTION AND SIGNAL TRANSDUCTION
Photoreception in somatic cells at the end of collagenic fiber optic pathways has not been demonstrated. However, many biomolecules absorb light at specific wavelengths and could function as cell photoreceptors for single photons. Low-intensity laser stimulation of neurons in Aplysia has been shown to induce membrane depolarization (Fork, 1971). Low-power laser stimulation of Auerbach's plexus in guinea pig ileum has been shown to increase acetylcholine release (Vizi et al., 1977) and low-power laser stimulation of the median nerve in humans has been shown to produce 'laser-induced somatosensory evoked potentials' in humans (Walker and Akhanjee, 1985). These reports suggest a general neural sensitivity to light exists at relatively high light intensities (though none were of such intensity to cause local tissue heating). However, more is known about vertebrate photoreceptors and their ciliary origin (see Figure 11 below) which relates to the possible role of endogenous photons in consciousness.
Figure 11: Specialized Cilia in Mammalian Photoreception
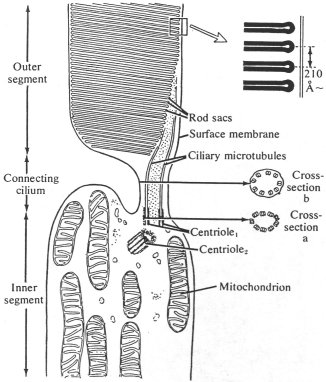
|
Diagram of a mammalian retinal rod cell, showing the centriole pair associated with the modified cilium of the outer segment. The outer segment contains rod sacs that are transversely arranged membranous disks formed from the folded ciliary membrane.
Adapted from Avers, 1976, p 387. |
The Role of Cilia in Sensory Reception
Retinal transduction of light into neural signals begins with the absorption of photons by the visual pigment rhodopsin, which is contained in a complex lamellar structure that is a highly modified cilium. Modified cilia also serve as signal transducers in the auditory, vestibular, and olfactory sensory systems. Ordinary cilia line airway surfaces, where their coordinated rhythmic beating moves a thin sheet of mucus constantly upward in the 'mucociliary escalator' so that adherent inhaled particles do not accumulate. Ordinary cilia are sometimes located in places where they have no known function, such as the glomerulus of the kidney (although one might speculate that they stir the glomerular filtrate, such stirring seems purposeless). Even highly modified ciliary photoreceptors resembling retinal photoreceptors are found where they have no known function: in the pineal organ of the brain. Cilia have clearly been multipurpose tools throughout evolution.
Evolution Builds Specialized Structures From Simpler Ones
A basic evolutionary principle is that complex structures are incrementally derived from more primitive structures having rudimentary capabilities of a similar nature. Even unicellular organisms may possess cilia, which often are used like little paddles for locomotion. But how did cilia evolve into specialized structures for photoreception? There had to have been many evolutionary intermediaries spanning the gap from simple to complex; is it possible that even the simplest cilia have rudimentary photoreceptive or waveguiding capabilities? Flagella are structurally very similar to cilia (both are made of microtubules in the same cross-sectional configuration) but they are longer and often occur in pairs. The unicellular alga Euglena Gracilis swims toward sunlight using the longer of its two flagella. Its shorter non-emergent flagellum may possibly be involved in photoreception as a 'comparator device' with the rhodopsin in the paraxial swelling (Insinna, in press) although the exact mechanisms of Euglena's phototaxis are unknown.
Figures 12 and 13: Ciliary Involvement in Primitive Photoreception
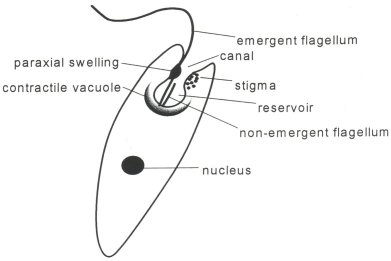
|
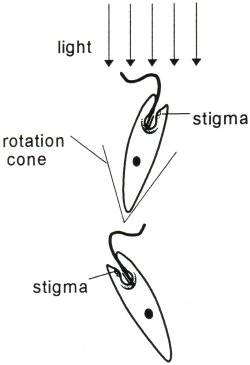
|
| Major organelles of Euglena Gracilis. Note the close association of the non-emergent flagellum with the paraxial swelling. | Euglena swims toward a light source as long as the light is within its rotation cone. It rotates approximately twice per second. It is believed (Insinna, in press) that the pigmented stigma casts a shadow on the rhodopsin-containing paraxial swelling to enable Euglena to orient to light. |
Adapted from Insinna (in press).
CELL INTELLIGENCE
Guenter Albrecht-Buehler has gathered very good evidence for physiologically functional endogenous light being functional in cell-to-cell communication. A summary of his work is at http://www.basic.northwestern.edu/g-buehler/cellint0.htm.
Albrecht-Buehler has shown that infrared light affects cell motility. But cells emit photons throughout the visible range and even the near ultraviolet; these endogenous photons have shorter wavelengths and greater energy than photons in the infrared range. If infrared photons are physiologically functional, it would therefore seem entirely possible that higher-energy photons would have physiological functions too. It may well be that light of different wavelengths has effects other than on cell motility, such that specific functions are mediated by certain wavelengths or ranges of wavelengths.
The important significance of intelligence expressed on a cellular level through endogenous photons would be its scaling up from individual cells to aggregations of cells through the range of multicellular organisms to the mammalian brain. Obviously more work in many fields will be needed if the hypothesized nexus is found.
SUMMARY
Cells are known to emit light, there is abundant historical reference to our spiritual nature being associated with light, and there are physiological structures and functions in the brain that could organize endogenous light into resonant interference patterns. Since interference patterns can themselves interfere with one another, endogenous light in the brain may produce a nexus through which a tiny portion of a broader substrate of interfering light in its own frame of reference (aka God) is enabled to experience an individualized consciousness. These mechanisms are consistent with a holographic model of the universe and some spiritually oriented views about the paradoxical unity of individual consciousness with God-consciousness:
"We are all part of the One Spirit. When you experience the true meaning of religion, which is to know God, you will realize that He is your Self, and that He exists equally and impartially in all beings."
Paramahansa Yogananda
ACKNOWLEDGEMENTS
This theoretical framework was first presented on Tuesday, April 11th, at a scientific meeting titled "Tucson 2000: Toward a Science of Consciousness" held April 10-15, 2000 at Tucson, Arizona, sponsored by the Center for Consciousness Studies at the University of Arizona. It is dedicated to the memory of the late Ray Burger, a friend and academic mentor who precipitated my development of this concept by enlightening me with the knowledge that the ventricles of the brain are lined with cilia, physiologic function unknown, when I already knew about endogenous light. Thank you, Ray.
GLOSSARY
- Bleb - a small piece or outpouching such as a blister or wart, usually with a connotation of maintaining contact with the larger body.
- Endogenous - originating or produced within an organism, tissue, or cell; in this context endogenous light is emitted from cell mitochondria.
- Ependyma or ependymal cells - layer of ciliated cells lining much of the inner ventricular surfaces and the central canal of the spinal cord.
- Explanatory Gap - refers to the failure of present knowledge in neuroanatomy and neurophysiology to explain consciousness; another way of referring to the same conundrum as the 'hard problem'.
- Hard Problem - how is it that we have subjective experience, and can experience experiencing? According to Chalmers:
The really hard problem of consciousness is the problem of experience. When we think and perceive, there is a whir of information-processing, but there is also a subjective aspect. As Nagel (1974) has put it, there is something it is like to be a conscious organism. This subjective aspect is experience. When we see, for example, we experience visual sensations: the felt quality of redness, the experience of dark and light, the quality of depth in a visual field. Other experiences go along with perception in different modalities: the sound of a clarinet, the smell of mothballs. Then there are bodily sensations, from pains to orgasms; mental images that are conjured up internally; the felt quality of emotion, and the experience of a stream of conscious thought. What unites all of these states is that there is something it is like to be in them. All of them are states of experience.
(see Chalmer's paper, Facing Up to the Problem of Consciousness for a fuller philosophical treatise on the 'hard problem').
REFERENCES
Chalmers, D. Facing up to the problem of consciousness. http://www.u.arizona.edu/~chalmers/papers/facing.html (undated, from an article by the same title in the Journal of Consciousness Studies, 1995).
Colli, L., Facchini, U., Guidotti, G., Lonati, R.D., Orsenigo, M., Sommariva, O. Further measurements on the bioluminescence of the seedlings. Experientia 11: 479-481, 1955.
Drollett, D. Vegetable lasers provide food for thought. Biophotonics International Jan/Feb p 32, 2000.
Fork, R.L. Laser stimulation of nerve cells in aplysia. Science 171:907-908, 1971.
Hameroff, S. Quantum computation in brain microtubules? The Penrose-Hameroff 'Orch OR' model of consciousness. Phil. Trans. R. Soc. Lond A356:1869-1896, 1998.
Insinna, E.M. Ciliated cell electrodynamics: from cilia and flagella to ciliated sensory systems. To be published in A. Malhotra (Ed.), Advances in Structural Biology v. 5, JAI Press Inc., Stamford, CT.
Jibu, M., Hagan, S. Hameroff, S.R., Pribram, K.H., Yasue, K. Quantum optical coherence in cytoskeletal microtubules: implications for brain function. Biosystems 32:195-209, 1994.
Johnson AR, Nixon RH, and Bergman LA. Optical Addressing and Clocking of RAM's. NASA Tech Briefs 13:32-34, 1989.
Llinas, R.R., Pare, D. Commentary: Of dreaming and wakefulness. Neuroscience 44:521-535, 1991.
Newman, J. Thalamic contributions to attention and consciousness. Consciousness and Cognition 4:172-193, 1995.
Ruth, B., Popp, F.A. Experimentelle untersuchungen zur ultraschwachen photonenemission biologischer systeme. Zeitschrift fur Naturforschung 31c:741-745, 1976.
Simanonok, K.E. Elastooptic photon signal modulation in collagenic fiber optics of tendon. Biomaterials, Medical Devices, and Artificial Organs 11:83-92, 1983.
Talbot, M. The Holographic Universe HarperCollins Publishers, Inc., New York, 1992
Tarusov, B.N., Polivoda, A.I., Zhuravlev, A.I. Study of the faint spontaneous luminescence of animal cells. Biophysics 6:83-85, 1961.
Veselovskii, V.A., Sekamova, Y.N., Tarusov, V.N. Mechanism of ultraweak spontaneous luminescence of organisms. Biophysics 8:147-150, 1963.
Vizi, E.S., Mester, E., Tisza, S., Mester, A. Acetylcholine releasing effect of laser irradiation on Auerbach's plexus in guinea-pig ileum. J. Neural Transmission 40:305-308, 1977.
Vladimirov, Y.A., L'vova, O.F. Superweak luminescence and oxidative phosphorylation in mitochondria. Biophysics 9:548-550, 1964.
Walker, J.B., Akhanjee, L.K. Laser-induced somatosensory evoked potentials: evidence of photosensitivity in peripheral nerves. Brain Research 344:281-285, 1985.
Wu, W.H., Bergman, L.A., Johnston, A.R., Guest, C.C., Esener, S.C., Yu, P.K.L., Feldman, M.R., Lee, S.H. Implementation of Optical Interconnections for VLSI. IEEE Transactions on Electron Devices ED-34:706-714, 1987.
Yamada, K.M., Olden, K., Hahn, L.E. Cell surface protein and cell interactions. In Subtelny, S., Wessells, N.K. (Eds.) The Cell Surface: Mediator of Developmental Processes Academic Press, 1980, 43-77.
Zhuravlev, A.I., Tsvylev, O.P., Zubkova, S.M. Spontaneous endogenous ultraweak luminescence of the mitochondria of the rat liver in conditions of normal metabolism. Biophysics 18:1101-1105, 1973.